The Ear
Anatomy
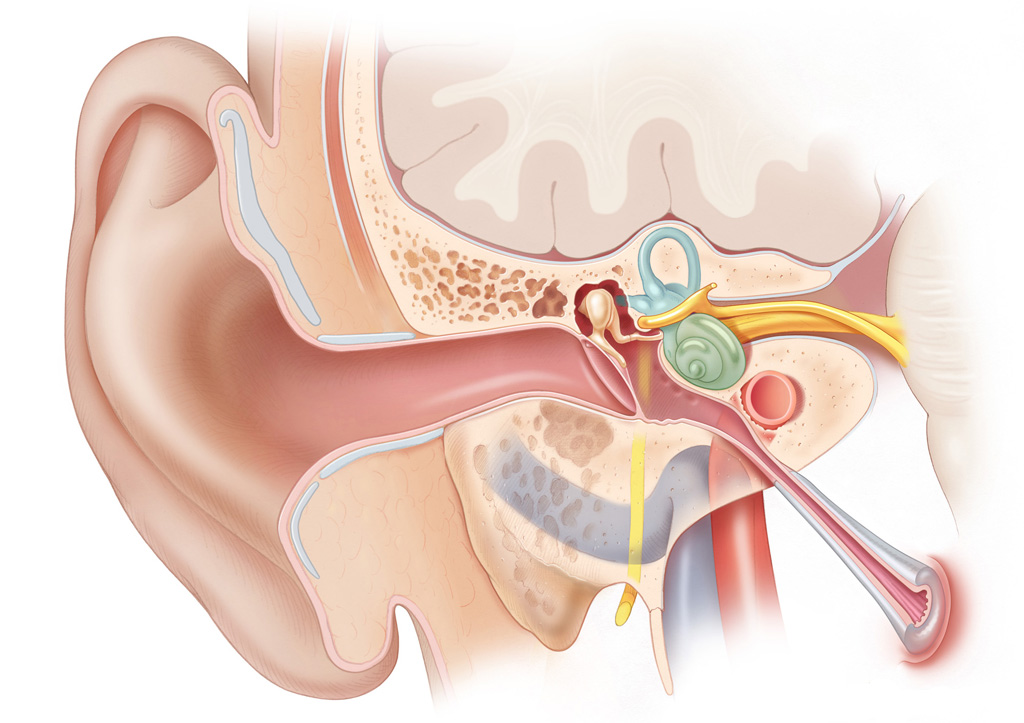
The peripheral auditory system has three main parts: the external ear, which includes the auricle and the external auditory canal (EAC), the middle ear, which includes the ossicles, and finally, the inner ear, which includes the cochlea. The peripheral vestibular system consists of the three semicircular canals, the utricle, and the saccule.
External Ear

The auricle or pinna refers to the external cartilaginous portion of the ear, which protrudes laterally from the side of the head. It is continuous with the EAC.
The EAC is approximately 2.5 cm in length. The lateral (outer) one-third of the EAC is cartilaginous and is referred to as the membranous EAC. This segment of the EAC is lined by thick, hair-bearing skin and cerumen producing glands. The medial (inner) two-thirds of the EAC is made up of the tympanic portion of the temporal bone and is lined by thin, hairless skin that is continuous with the tympanic membrane.
The tympanic membrane separates the external auditory canal from the middle ear space. It consists of an outer layer of squamous epithelium, a middle fibrous layer, and an inner layer of mucosal epithelium. The majority of the outer circumference of the tympanic membrane is anchored to the tympanic sulcus of the bony EAC via a fibrous annulus.
lateral process of the malleus
The superior portion of this annulus is incomplete; at the termination of the tympanic sulcus, the fibrous tissue forms the anterior and posterior malleal folds, which attach to the. The manubrium of the malleus then extends anteroinferiorly, tenting the tympanic membrane outward. The tip of the malleus attaches to the tympanic membrane at the umbo. There are two distinct regions of the tympanic membrane: the pars tensa and the pars flaccida. The pars tensa lies inferior to the anterior and posterior malleal folds. The pars flaccida is the portion of the tympanic membrane superior to the anterior and posterior malleal folds; it is so named because it has a weaker middle fibrous layer.
Blood Supply and Innervation
The posterior auricular and superficial temporal arteries (branches of the external carotid artery) provide blood supply to the external ear. Sensation to the auricle and EAC is provided by the great auricular nerve (from C2 and C3 nerve roots), the auriculotemporal branch of cranial nerve V, and a cutaneous branch of cranial nerve VII.
Middle Ear
The middle ear space contains the ossicles, or bones of the ear, and is divided into three regions: the hypotympanum, mesotympanum, and epitympanum. The mesotympanum is located directly behind (medial to) the tympanic membrane. The opening of the Eustachian tube is located in the anterior region of the mesotympanum. Posteriorly, the mesotympanum opens into the sinus tympani. The mesotympanum ends at the inferior limit of the tympanic annulus; inferior to this is the hypotympanum, which extends to the jugular bulb. Superiorly, the mesotympanum ends at the lateral process of the malleus, and the epitympanum begins. The epitympanum contains the bulk of the malleus and incus, which are suspended within the epitympanic space by ligaments. The scutum refers to the superomedial wall of the bony canal, which forms the lateral wall of the epitympanum. Posteriorly, the epitympanum communicates with the antrum of the mastoid (see the section on temporal bone anatomy, below). Superiorly, the epitympanum ends at the tegmen, which is the floor of the middle fossa of the cranium.
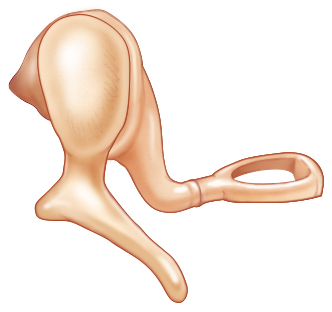
There are three ossicles: the malleus, incus, and stapes. The malleus consists of a manubrium (or long process, which is attached to the tympanic membrane), a lateral process, and a head. Within the epitympanum, the head of the malleus articulates with the body of the incus at the incudomalleolar joint. The incus has a long process and a short process. The short process is attached to the posterior wall of the middle ear at the fossa incudis of the mastoid bone. The long process articulates with the stapes at the incudostapedial joint. The stapes consists of the superstructure, which is a wishbone-shaped component, and the footplate. The stapes superstructure consists of the anterior and posterior crura, which converge to form the capitulum, which in turn articulates with the incus at the lenticular process. The footplate of the stapes sits within the oval window.
The Eustachian tube connects the middle ear space to the nasopharynx. The proximal one-third of the Eustachian tube is bony, while the distal two-thirds is cartilaginous. In the adult, the Eustachian tube travels inferomedially at an approximately 45 degree angle, ending at the torus tubarius in the nasopharynx.
The tensor veli palatini muscle inserts laterally on the Eustachian tube, opening the tube during actions such as yawning and swallowing. The two major functions of the Eustachian tube are ventilation and drainage of middle ear secretions into the nasopharynx.
Inner Ear
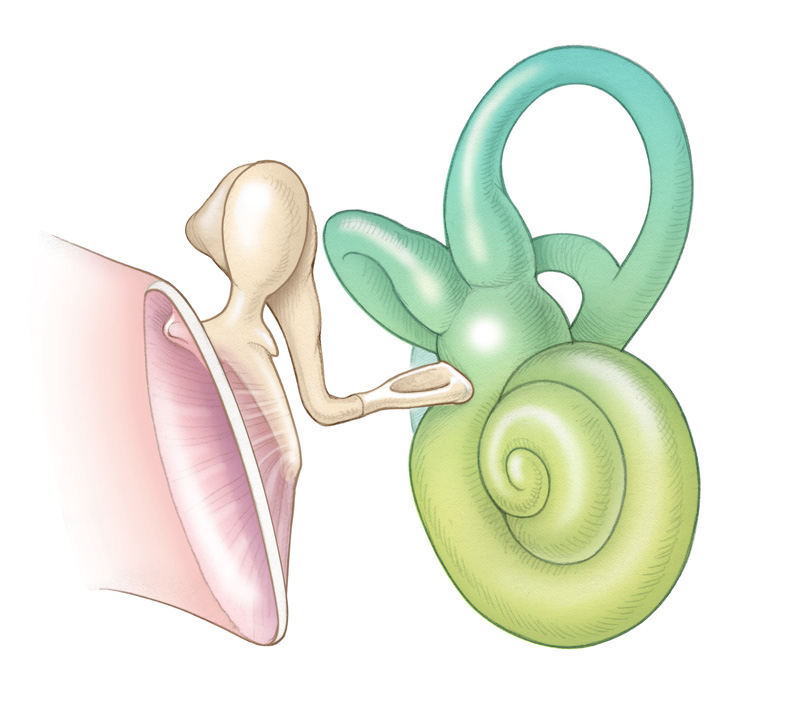
The inner ear, which consists of the cochlea, utricle, saccule, and semicircular canals, is located in the temporal bone . The structures of the inner ear can be thought of as two sets of telescoping tubes, one inside the other. The outer tube is osseous and therefore referred to as the bony labyrinth. It is filled with a fluid called perilymph, which is similar in composition to cerebrospinal fluid. Within the bony labyrinth runs the second set of tubes, called the membranous labyrinth. The membranous labyrinth is filled with a fluid called endolymph, which is similar in composition to intracellular fluid.
At the boundary of the middle ear, the stapes footplate contacts the oval window membrane, which transmits vibratory energy into the central section of the labyrinth, the vestibule. The vestibule is continuous with both the semicircular canals and the cochlea. The cochlea is a snail-shaped structure with a wide base and a narrow apex. The bony labyrinth includes the otic capsule (bony border of the cochlea) and modiolus, the bony tube of the cochlea, where fibers of the auditory nerve enter from the internal auditory meatus to synapse onto hair cells of the inner ear. As the modiolus extends from base to apex, the interior bony labyrinth is partitioned into three separate scalae (chambers): the scala vestibuli, scala tympani and the scala media. The most apical scala, the scala vestibuli, is partitioned from the scala media by Reissner’s membrane. The scala media is partitioned from the scala tympani by the osseous spiral lamina and basilar membrane. Both the scala vestibuli and the scala tympani contain perilymph and are connected at the most apical region of the cochlea, a region called the helicotrema, where perilymph flows from one scala to the other. Proximally, the scala vestibule ends at the oval window and the scala tympani at the round window.
The scala media houses the cells responsible for the perception of sound. The organ of Corti refers to a complex set of structures and cell types situated on the basilar membrane that make up the sensory organ of hearing. Medially, the organ of Corti contains one row of inner hair cells, while laterally there are three rows of outer hair cells. Between the inner and outer hair cells are the pillars of Corti. Overlying the inner and outer hair cells is a gelatinous sheet of collagen fibers called the tectorial membrane. Each hair cell contains a thickened region at its apex called the cuticular plate. From the cuticular plate, bundles of mechanoreceptors called stereocilia project outward (thus the apt name of ‘hair cells’). The stereocilia are arranged in order, from longest to shortest, and are connected to one another so that each bundle moves together as one unit. The longest stereocilia of the outer hair cells contacts the overlying tectorial membrane, allowing them to deflect with transmission of sound waves. Details regarding the physiology of the auditory system are discussed below.
The vestibular organs of the inner ear comprise the three semicircular canals, which detect angular acceleration, and the two otolithic organs (the utricle and saccule), which detect linear acceleration. The semicircular canals are paired such that each canal on one side of the head lies in the same plane as another canal on the opposite side. The two horizontal (sometimes referred to as lateral) canals are in the same plane; the right posterior canal is in the same plane as the left superior (sometimes referred to as anterior) canal, and vice versa. Each semicircular canal has a dilated end called the ampulla that empties into the utricle, which is located in the vestibule, along with the saccule. The nonampullated ends of the posterior and superior canals merge to form a common crus. The ampulla of each semicircular canal contains the crista ampullaris, which is the neuroepithelial structure that bears the receptor hair cells of the vestibular system (akin to the organ of Corti in the cochlea). The stereocilia of the vestibular hair cells project outward from the crista into a gelatinous structure called the cupula. Similarly, the surfaces of the utricle and saccule are covered with hair cells. Their stereocilia protrude outward and are embedded in the gelatinous otolithic membrane, which also contains calcium carbonate crystals called otoconia (Figure 1.10). The endolymphatic duct is a canal that extends from the anterior aspect of the utricle to end in the endolymphatic sac, which is in contact with the dura. Another, smaller canal (the ductus reuniens) connects the saccule to the cochlea. Details regarding the physiology of the vestibular system are discussed below.
Temporal Bone
The temporal bone is a pyramidal-shaped bone with four embryologically distinct components: squamous, mastoid, petrous, and tympanic . The squamous portion of the temporal bone forms part of the middle fossa and contains the insertions of the temporalis and masseter muscles. The mastoid portion of the temporal bone is pneumatized and aerated, consisting of air-filled cells. The central air cell is called the antrum. The inferior portion of the mastoid is elongated to form the mastoid tip, onto which the sternocleidomastoid muscle inserts. The posterior belly of the digastric muscle inserts in a sulcus medial to the mastoid tip. The petrous portion of the temporal bone contributes to the floor of the middle fossa and forms the anterior boundary of the posterior fossa. The entrance of the internal auditory canal (porus acusticus) and the foramina for cranial nerves VII and VIII are located in the posterior surface of the petrous portion of the temporal bone. Finally, the tympanic portion of the temporal bone contributes to the bony external auditory canal and the middle ear.
Facial Nerve
Understanding the course of the facial nerve (cranial nerve VII) is essential for the surgeon operating in the ear and skull base. Fibers of the facial nerve originate from the motor nucleus, superior salivatory nucleus, and nucleus of the solitary tract in the brainstem. The nerve emerges from the brainstem just superior to the cerebellopontine angle, enters the temporal bone at the porus acusticus of the internal auditory canal and travels within the fallopian canal until it reaches the stylomastoid foramen, where it branches out into five main divisions. The first portion of the fallopian canal is referred to as the labyrinthine segment. This is the most narrow segment and ends at the geniculate ganglion. Here, the greater superficial petrosal nerve branches off to provide parasympathetic innervation to the lacrimal gland and sensory taste fibers to the palate. The facial nerve makes an acute turn at the first genu to enter the second segment of the fallopian canal, called the tympanic or horizontal segment. At the second genu, the facial nerve turns inferiorly to become the mastoid or vertical segment. At variable points along this last segment of the fallopian canal, the chorda tympani and nerve to stapedius branch off. The facial recess is a region in this area, bounded by the facial nerve, the chorda tympani, and the fossa incudis. Access to the middle ear is gained through this recess when approaching from the mastoid portion of the temporal bone (e.g., for performing cochlear implant insertion). The facial nerve exits the temporal bone to enter the face at the stylomastoid foramen.
Physiology
Auditory System
The perception of sound begins with the generation of mechanical vibratory sound waves by a stimulus. These vibratory waves are propagated through the air until they reach the ear. The pinna serves as a funnel, directing sound waves into the external auditory canal, as well as providing a small degree of sound amplification. At the termination of the EAC, the sound waves cause vibration of the tympanic membrane, which then results in vibrations of the ossicles. Sound is then transmitted into the inner ear as the stapes footplate vibrates the oval window. At this point, the sound waves transition from travel in an air-filled environment to a fluid-filled chamber.
Within the inner ear, transmitted compression waves travel through the cochlea toward the round window. As the fluid wave travels across the compartments of the cochlea, a pressure gradient is created between the scala vestibuli and scala tympani. This causes a vibration of the basilar membrane, producing a traveling wave that moves from the base of the cochlea to the apex. Because the basilar membrane decreases in stiffness across its length, with the highest stiffness at the base and lowest at the apex, sounds of differing frequencies resonate at different places along the basilar membrane. This constitutes the tonotopic organization of the cochlea.
The traveling wave created across the basilar membrane causes the inner hair cell stereocilia to shear relative to the tectorial membrane. The stereocilia are then deflected, causing stretching of the filamentous tip links between the rows of stereocilia. This results in the opening of stretch-sensitive ion channels and depolarization. At this point, the mechanical (vibratory) energy of the sound wave is transduced into electrical energy. Like the basilar membrane, the hair cells are also tonotopically arranged, such that each hair cell has a characteristic frequency to which it is most sensitive.
Depolarization of the inner hair cells results in the opening of voltage-gated calcium channels, which then triggers the release of neurotransmitters across the synapse with auditory nerve fibers. From here, the electrochemical signal is propagated along the auditory nerve. The afferent auditory neurons are called spiral ganglion cells, and consist of two types. Type I spiral ganglion cells predominate (90%); each of these myelinated cells forms a synapse with a single inner hair cell. Each inner hair cell, in turn, may have synapses with multiple type I spiral ganglion cells. Type II spiral ganglion cells (10%) are unmyelinated and form synapses with multiple outer hair cells.
The auditory nerve fibers terminate on cells in the cochlear nucleus of the brainstem at the pontomedullary junction. The cochlear nucleus consists of several subdivisions, each containing specific cell types with different targets. These subdivisions are responsible for identifying the type, location, and intensity of the sound stimulus. Projections from the cochlear nucleus are sent to the superior olivary complex, lateral lemniscus, and inferior colliculus, which are additional brainstem structures that continue processing and integrating the sound information. From the brainstem, the auditory information travels to the medial geniculate body of the thalamus, which is thought to further refine sound localization as well as process complex sound patterns, such as human speech. Finally, the auditory input is transmitted to the primary and association auditory cortices4 of the cerebral cortex, located in the temporal lobe. Like the cochlea, the primary auditory cortex is also tonotopically organized.
Vestibular System
The primary function of the vestibular system is to assist in balance and orientation by sensing and processing information regarding the body’s motion. By detecting the position of the head relative to gravity, the vestibular system can maintain visual and postural stability through activation of compensatory movements. The semicircular canals are responsible for detecting rotational acceleration of the head. When the head rotates about the axis of a semicircular canal (SCC), movement of the endolymph within the canal causes deflection of the gelatinous cupula. This then deflects the stereocilia underlying the cupula on the crista ampullaris. Because the stereocilia are polarized, they are maximally stimulated by rotation in a particular direction. The presence of three semicircular canals therefore allows for detection of rotation in each of the three dimensional planes (x-, y-, and z-axis). The mechanical deflection of the hair cells in the SCC is transduced into an electrical signal by the activation of ion channels and subsequent neurotransmitter release from the hair cell membrane. This produces a change in the tonic firing rate of vestibular afferent nerve fibers, either inhibiting or increasing the firing rate, depending on the direction of hair cell deflection. This information is transmitted along the afferent nerve fibers to secondary vestibular neurons in the brainstem, which then project to the ocular motor nuclei, spinal motor neurons, autonomic centers, cerebellum, and cerebral cortex. This allows for the generation of reflexive responses to counteract the movement of the head. For example, stimulation of a semicircular canal produces eye movements in the plane of that canal. In cases of vestibular dysfunction where there is an imbalance between the inputs from the left and right canals, the resulting reactive eye movements are conflicting, producing the phenomenon known as nystagmus.
The utricle and saccule function in a similar manner to detect linear acceleration of the head. The utricle is responsible for detecting acceleration in a horizontal (axial) plane, while the saccule detects acceleration in a vertical (parasagittal) plane. Linear acceleration of the head produces a shear force between the gelatinous otolithic membrane9 and the underlying macula (analogous to the cupula and crista of the SCCs). This then results in hair cell deflection, signal transduction, and modulation of afferent nerve fiber tonic firing rate, much in the same manner as in the SCCs. Similarly, compensatory reflexes also occur in response to linear acceleration. For example, saccular excitation is interpreted by the body as falling, and results in activation of the extensor muscles of the trunk and limb in an effort to restore postural stability. Additional details regarding the vestibular system can be found in Vestibular Disorders.